")


Dear visitor,
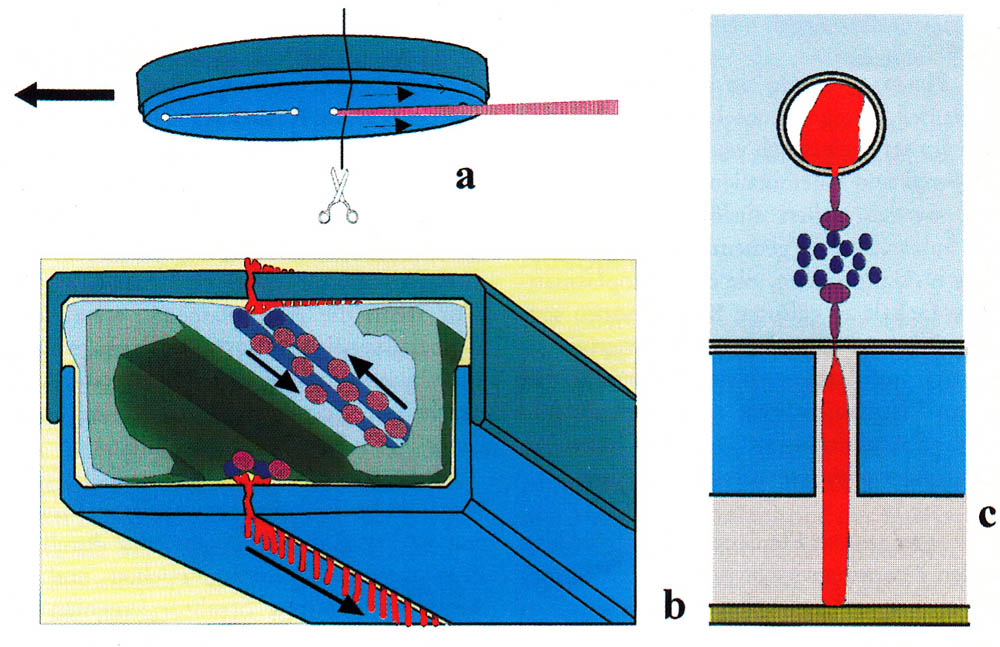
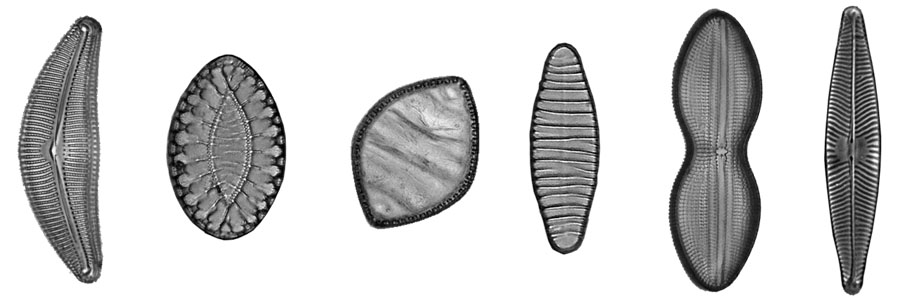
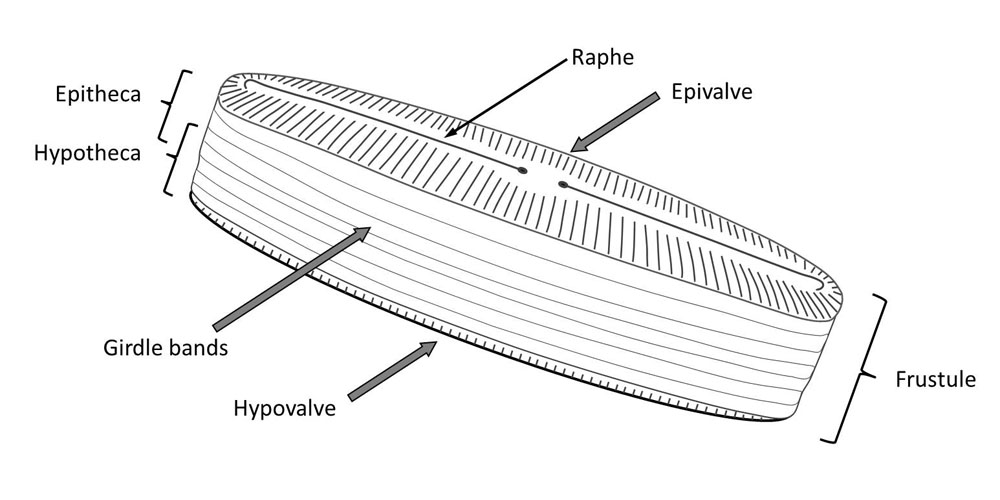
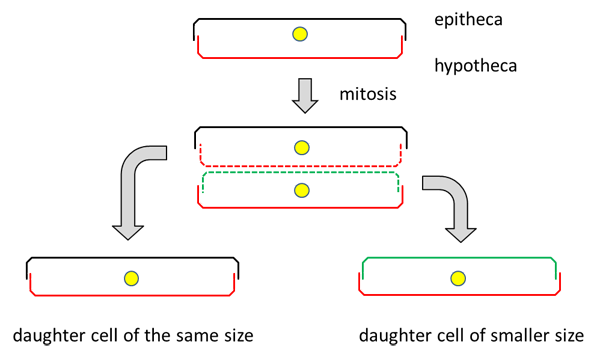
You may already have seen images of diatoms with their diverse forms and fine structures. Perhaps you also had the opportunity to observe live diatoms under the microscope and noticed that some are capable of moving smoothly or jerky. They often change the direction of movement. The mechanism of movement is not visible.
One of my favourite pastimes is observing diatoms with the focus on species that are motile. I am fascinated by the reaction of diatoms to environmental conditions and external stimuli, such as obstacles or light. In cultures of colonial diatoms, I try to understand the link between motility and the formation of colonies.
I would like to mention that I (Thomas Harbich, see contact) studied physics and spent my working life with typical activities of an engineer in the field of telecommunications and the automotive area. Since a few years I am retired.
The purpose of this homepage is to encourage you to make your own observations on diatoms. If you like to microscope and follow this suggestion, you will hopefully find useful information on these pages.
You will get information about the following aspects:
- Cultivation of diatoms
- Observation and creation of video recordings of the movement of diatoms in different environments
- Analysis of the resulting images and videos
- Hypotheses and conclusions from the observations
As there is a comprehensive literature and many web pages to the biology of the diatoms, I restrict myself to some remarks, links and literature references.
From time to time the site will be supplemented by observations and hypotheses. If you are interested in the subject, please feel invited to visit this website now and then. I am grateful for content discussion and suggestions for improvement.
Please note that the content is protected by copyright. If you quote something from this page, please refer to my name and add a link to this page. Please contact me if you want to make use of pictures or videos. I do not pursue any commercial interests.
The site contains many short videos. To view them, a browser is required, which masters HTML5. If a video on a browser is not running correctly, it is recommended to try another browser.
As an introduction to the topic, I show a picture from the book "Kunstformen der Natur" by Ernst Heinrich Philipp August Haeckel (16.02.1834 to 09.08.1919):
 |
 |

 and Applications) Vadim V. Annenkov (Editor), Richard Gordon (Series Editor), Joseph Seckbach (Series Editor), Wiley-Scrivener; First published: 29 October 2021,
and Applications) Vadim V. Annenkov (Editor), Richard Gordon (Series Editor), Joseph Seckbach (Series Editor), Wiley-Scrivener; First published: 29 October 2021,