
|
|
|
   |
   |
|
Eunotia sp. (300-facher Zeitraffer) und überlagerte Bilder |
Eunotia sp. (150-facher Zeitraffer) und überlagerte Bilder |
Die Bewegung von Eunotia sp.
Wie die bisherigen Beispiele gezeigt haben, lassen sich unterschiedliche Eigenschaften der Bahnen von Diatomeen auf glattem Substrat vor dem Hintergrund der Form ihrer Raphe verstehen. Dazu gehören Bahnkrümmung, Lage des Zentrums der Kraft und das Verhalten der Krümmung an Umkehrpunkten.
Nicht immer ergeben sich jedoch einfach interpretierbare Trajektorien. Die Bewegung bei Cymatopleura elliptica zeigt (siehe Video), dass bestimmte Formen von Valven in Zusammenhang mit der Form der Raphen nur eine zufällige Bewegung ermöglichen. Hier soll auf Eunotia eingegangen werden. Die Bewegung von Diatomeen dieser Gattung ist selbst in der Ebene, also ohne Aufrichten, ungewöhnlich.
Spezies und Raphen
Im März 2018 wurde in einem Teich bei Stuttgart/Hohenheim (48°42'34.0"N 9°12'29.1"E) eine Eunotia sp. isoliert und klonal kultiviert. Ich gehe davon aus, dass es sich um Eunotia glacialis Meister 1912 handelt. Angesichts der vorhandenen Bilder könnte es sich nach Hinweisen von Frau Dr. Nélida Abarca (Botanischer Garten und Botanisches Museum Berlin, Freie Universität Berlin) auch um Eunotia glacialifalsa, Eunotia glacialispinosa, Eunotia belgica oder Eunotia valida handeln. Entsprechend der dokumentierten Verbreitungsgebiete der genannten Spezies in Baden-Württemberg (Mattern et al. 2019) scheint die Zuordnung zu Eunotia glacialis als

 wahrscheinlich. Da eine Restunsicherheit bleibt, wird sie hier als Eunotia sp. bezeichnet. Die Bilder links zeigen die Gürtelband- und Valvenansicht. Die Diatomeen sind gerade aus einer Zellteilung hervorgegangen und deshalb ist ihre Breite (Ausdehnung in Richtung der Pervalvarachse) gering.
wahrscheinlich. Da eine Restunsicherheit bleibt, wird sie hier als Eunotia sp. bezeichnet. Die Bilder links zeigen die Gürtelband- und Valvenansicht. Die Diatomeen sind gerade aus einer Zellteilung hervorgegangen und deshalb ist ihre Breite (Ausdehnung in Richtung der Pervalvarachse) gering.
Bei der Gürtelbandansicht wird die kurze Raphe, soweit sie deutlich erkennbar ist, periodisch als gelbe Linie eingeblendet. Das Bild der Valvenansicht legt nahe, dass die Raphe an den Polen noch ein Stück auf der Valvenoberseite verläuft. Solche Raphen-Äste sind typisch für Eunotia.
In der Regel liegen diese Diatomeen mit den Gürtelbändern auf dem Substrat. Offenbar ist zumindest in Kultur meist die konkave Seite dem Substrat zugewandt. In dieser Lage können bis zu vier Raphen-Äste Kontakt zum Substrat besitzen und zum Antrieb beitragen. Die Voraussetzungen für die bereits beschriebenen Analyseverfahren sind hier offenbar nicht erfüllt.
Motilität
Nachfolgend wird nur die Bewegung in der Ebene in Gürtelbandlage betrachtet. Die von Bertrand 1993 beschriebenen Bewegungsmuster von Eunotia pectinalis zeigen eindrucksvoll, dass die Raphensysteme dieser Spezies auch vielfältige Bewegungen ermöglichen, bei denen sich die Diatomee aufrichtet. Solche Abläufe konnte ich nicht beobachten. Ein Kippen in die Valvenlage geschah bei der beobachteten Spezies in Kultur nur als Folge von Kollisionen zwischen Diatomeen.
   |
Die kultivierte Eunotia bildet kettenförmige Kolonien, die in vitro sehr lang werden können. Einige Bilder findet man auf den Seiten zur Größenfolge und Synchronizität kettenförmiger Kolonien. Beweglich sind die einzelnen Diatomeen, die sich von Kolonien abgelöst haben. Nur selten beobachtet man mehrere verbundene Diatomeen in Bewegung. Deshalb können sie wohl auch epiphytisch besiedelt werden. In der ersten Rohkultur waren neben Eunotia auch Cymbella enthalten, die Eunotia-Kolonien besiedelten. Das Video links (4-facher Zeitraffer) zeigt die Situation.
In den beiden Videos am Anfang dieser Seite (300-facher und 150-facher Zeitraffer) sieht man einige Diatomeen in Bewegung. Darunter sind jeweils in einer Bildergalerie Überlagerungsbilder von Frames in unterschiedlichen zeitlichen Abständen abgebildet. Das zweite Video links zeigt die Bewegung in höherer Vergrößerung und 20-fachem Zeitraffer. Darunter sind wieder verschiedene Überlagerungen abgebildet.
Im Zeitraffer wirkt die Bewegung unregelmäßig. Die Bewegungsrichtung und Ausrichtung der Zelle ändern sich häufig. Zur Messung der Geschwindigkeit wurde deshalb die Bewegung des Mittelpunktes der Diatomeen verwendet. Die Geschwindigkeit ist relativ gering. Sie lag bei den durchgeführten Messungen zwischen 1,1 µm/s und 2,3 µm/s. Bei einer Länge von etwa 70 µ von Apex zu Apex benötigt die Diatomee also typisch zwischen einer halben und einer Minute, um die eigene Zelllänge zurückzulegen.
Bewegungsformen
Die Bewegungsformen sind durch die Raphen und deren Aktivität geprägt. Bei Eunotia in Gürtelbandlage sind sie vielfältig, da vier unabhängige Raphenäste gleichzeitig in Kontakt mit dem Substrat stehen können. Wie die 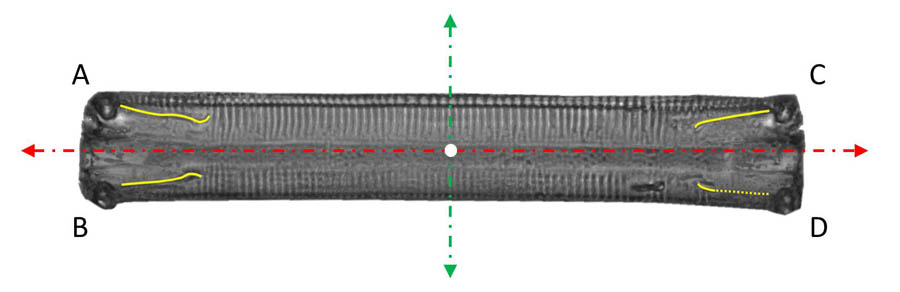 Videos der Bewegung und die überlagerten Bilder zeigen, bilden die Apikalachse (gestrichelt, rot im Bild links) und die Trajektorie des Mittelpunktes (weiß markiert) der Diatomee zumeist einen deutlich sichtbaren Winkel, d.h. die Ausrichtung der Diatomee ist nur selten tangential zur Bewegung des Mittelpunktes. Neben der Geschwindigkeitskomponente in Richtung der Apikalachse gibt es also meist eine wesentliche Komponente in Richtung der Pervalvarachse. Häufig erfolgt die Bewegung grob in der Richtung der Diagonalen der rechteckigen Umrisslinie (in Richtung der Strecken AD oder BC). Dies spricht dafür, dass dabei gegenüberliegende Raphenäste (z.B. die Raphenäste bei A und D) hauptsächlich zum Antrieb beitragen und in gleicher Richtung aktiv sind.
Videos der Bewegung und die überlagerten Bilder zeigen, bilden die Apikalachse (gestrichelt, rot im Bild links) und die Trajektorie des Mittelpunktes (weiß markiert) der Diatomee zumeist einen deutlich sichtbaren Winkel, d.h. die Ausrichtung der Diatomee ist nur selten tangential zur Bewegung des Mittelpunktes. Neben der Geschwindigkeitskomponente in Richtung der Apikalachse gibt es also meist eine wesentliche Komponente in Richtung der Pervalvarachse. Häufig erfolgt die Bewegung grob in der Richtung der Diagonalen der rechteckigen Umrisslinie (in Richtung der Strecken AD oder BC). Dies spricht dafür, dass dabei gegenüberliegende Raphenäste (z.B. die Raphenäste bei A und D) hauptsächlich zum Antrieb beitragen und in gleicher Richtung aktiv sind.
Gelegentlich wird die Komponente in apikaler Richtung sehr klein oder verschwindet und die Diatomee bewegt sich in Richtung der Pervalvarachse (gestrichelt, grün). Dies lässt sich durch eine gleichgerichtete Aktivität der Raphenäste etwa bei A und C erklären.
Große Bewegungskomponenten in Richtung der Pervalvarachse verleihen der Bewegung der Diatomeen in Zeitrafferaufnahmen etwas Schwebendes. Sie scheinen zu driften, obwohl sie eine starke Kohäsion an das Substrat besitzen.
Drehbewegungen
   |
Drehbewegungen sind ebenfalls zu beobachten. Das Video links zeigt eine Drehung um einen Vollkreis. Unterhalb des Videos sind Überlagerungen von Bildern zu sehen, die den gleichen zeitlichen Abstand besitzen.
Neben der hohen Regelmäßigkeit fällt auf, dass der Drehpunkt ein wenig außerhalb der Diatomee liegt. Eine naheliegende Erklärung der Drehbewegung besteht darin, dass nur die beiden Raphenäste A und B (bzw. C und D) Kontakt zum Substrat besitzen und ihre Aktivität gegensinnig verläuft. Eine Raphe arbeitet in apikale, die andere in distale Richtung. Dies ist im nachfolgenden Bild skizziert.

In dem konkreten Fall dauerte eine Drehung um 2π ziemlich genau 60 Sekunden, was eine Winkelgeschwindigkeit von ω = 0,105 rad/s bedeutet. Da die Breite der Diatomee bei 24,8 µm liegt, beträgt der Radius des Kreises etwa r = 12,4 µm. Mit der Beziehung v=r*ω erhält man eine Geschwindigkeit an der Raphe von 1,26 µm/s. Dies entspricht der typischen Geschwindigkeit dieser Spezies.
Bewegungsabläufe
Die gegebene Beschreibung ist grob vereinfacht, denn die Bewegungsrichtungen wechseln laufend. Drehungen und diagonale lineare Bewegungen überlagern sich häufig, so dass sich auf superponierten Bildern fächerartige Strukturen zeigen. Die Ausrichtung der Apikalachse ändert sich andererseits nicht zwingend, wenn die Trajektorie des Mittelpunktes eine Richtungsänderung vornimmt.
Bei vielen motilen Diatomeen kann man beobachten, dass längere Bahnen durchlaufen werden und plötzlich eine Richtungsumkehr erfolgt. Hier befindet sich meist ein Raphenast im Kontakt mit dem Substrat, dessen Aktivitätsrichtung sich ändert. In dieser deutlichen Form ist das bei der beobachteten Eunotia nicht zu erkennen. Lediglich ein Hin- und Herruckeln, bei dem eine Richtungsänderung schwer erkennbar ist, tritt auf. Eine mögliche Erklärung ist, dass bei Eunotia bis zu vier Raphenäste zur Bewegung beitragen können und es sehr wahrscheinlich ist, dass mehr als ein Raphenast zur Bewegung beiträgt. Dann führt die Umkehr der Bewegungsrichtung eines Raphenastes zwar zu einer Änderung der Richtung, nicht aber zu einer Umkehr der Diatomee.
Hinweis
Zu dem Thema der Bewegung von Diatomeen der Gattung Eunotia Ehrenberg hat Paula Furey folgendes Paper veröffentlicht, das auch Bezug auf diese Seite nimmt:
Furey, Paula C. "Motility in the Diatom Genus Eunotia Ehrenb." Diatom Gliding Motility (2021): 185-209.
Bertrand, J. (1993) Mouvements des diatomées. III. Le pivotement polaire vertical de Eunotia pectinalis (Kütz) Rab. Essai de quantification des forces. Cryptogam. Algol. 14(4), 157-172.
Mattern, H., Stutz, S., van de Vijver, B. und King, L. (2019). Klasse Bacillariophyceae. In: Beiträge zu den Algen Baden - Württembergs. Band 2: Spezieller Teil: Euglenozoa und Heterkontobionta (Ed. by S. Stutz & H. Mattern), pp. 94‒371. Verlag Manfred Hennecke, Remshalden.